1.B.34 The Corynebacterial Porin A (PorA) Family
The mycolata are a group of mycolic acid-containing bacteria which include the genera Rhodococcus, Gordona, Dietzia, Tsukamurella, Corynebacterium, Mycobacterium and Norcadia. They have an outer membrane in which the constituent mycolic acids are linked via ester bonds to the arabinogalactan that is attached to the murein of the cell wall. The mycolic acids are 2-branched, 3-hydroxylated fatty acids of varying lengths depending on the organism (i.e., 60-90 carbon atoms for the Mycobacteria, 46-58 carbon atoms for the Norcadia and 22-38 carbon atoms for the Corynebacteria). Porins allow permeation of small hydrophilic molecules across the outer membrane permeability barrier (see the MBP family, TC# 1.B.24).
PorA is a small (45 aa) porin of Corynebacterium glutamicum with an excess of four negative charges in agreement with its cation selectivity. It forms wide, oligomeric water-filled pores. It is encoded by a 138 bp gene, porA. Deletion of this gene from C. glutamicum results in slow growth and lower antibiotic sensitivities. PorA is therefore believed to be a major hydrophilic pathway through the cell wall (outer membrane) of some Corynebacteria (Costa-Riu et al., 2003).
PorA does not have an N-terminal leader sequence suggesting that its export across the cytoplasmic membrane does not occur via the Sec pathway. Only Corynebacteria possess homologues of PorA. C. glutamicum has only one gene coding for a PorA homologue. C. efficiens does not have a porA gene in its genome, so another outer membrane porin must serve the equivalent of the PorA function. Both C. glutamicum and C. efficiens have two homologous anion-selective outer membrane porins called PorB and PorC. They are in the PorB family (TC #1.B.41) and provide outer membrane, anion selective pore functions (Costa-Riu et al., 2003).
C. diphtheriae does have a 43aa PorA. It lacks a signal sequence but shows 40% identity and 62% similarity (with no gaps) in a stretch of 26 compared residues (15-41 in the C. glutamicum PorA). The C. diphtheriae PorA had an apparent molecular mass of about 66 kDa as determined on Tricine-containing sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels and consisted of subunits having a molecular mass of about 5 kDa. Single-channel experiments with the purified protein suggested that the protein formed channels with a single-channel conductance of 2.25 nS in 1 M KCl. Further single-channel analysis suggested that the cell wall channel is wide and water filled because it has only slight selectivity for cations over anions and its conductance followed the mobility sequence of cations and anions in the aqueous phase (Schiffler et al., 2007).
More recent studies revealed that PorA and PorH (1.B.59) form heterooligomeric pores, and in spite of the earlier reports, they are both required for pore formation (Barth et al. 2010). They comprise the major outer membrane porin in these Corynebacteria, and no other protein is required. However, they are apparently post-translationally modified by mycolic acids (Rath et al. 2011). Although the molecular weight of each subunit is about 5 KDa, the complex has an apparent molecular mass of about 66 KDa, suggesting that it contains about 12 subunits (Schiffler et al. 2007). Porin A-Porin H ion channels have been reconstituted in liposomes. With properly mycoloylated proteins, it manifests the typical voltage dependent ion channel properties of an outer membrane porin (Hünten et al. 2005; Rath et al. 2013).
PorA and PorH are of about the same size and topology, show substantial sequence similarity with each other, and are homologous (T. Su and MH Saier, unpublished observations). In contrast to most outer membrance proteins in Corynebacteria which have signal sequences and beta-barrel structures resembling those of Gram-negative bacteria (Marchand et al. 2012), these small proteins seem to lack signal sequences, and may have strongly amphipathic transmembrane alpha-helical structures (T. Su and MH Saier, unpublished results).
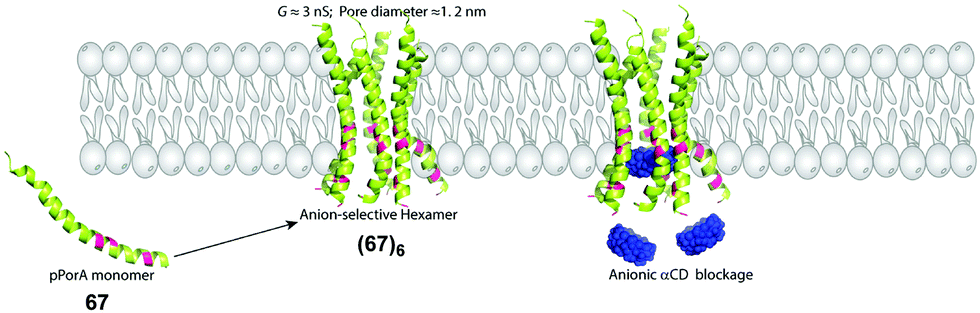
Mahendran designed an anion selective pore using synthetic peptides that derive from the protein PorACj from Corynebacterium jeikeium (see figure below). The wild-type pPorA monomer 67, consisting of 40 amino acids, folds into an α-helix and self-assembles in the membrane to give a highly conducting pore (G = 3.0 ± 0.2 nS at +50 mV) that is anion-selective (PCl−/PK+ ∼ 3/1). They proposed that pPorA 67 forms a hexameric pore (67)6, with diameter = 1.2 nm (Davis et al. 2020). Two experiments were consistent with this proposal that positioning of cationic and anionic sidechains at the mouth of the pore controls ion selectivity. First, conductance was abruptly and completely turned off upon addition of anionic α-cyclodextrins. This result is consistent with these anionic sugars being electrostatically attracted to the mouth of the pore and then getting lodged in the lumen, thus blocking ion transport. Second, both the size (∼1.4 nm) and ion selectivity of the channel were significantly altered by a single point mutation that changed a cationic lysine to a neutral cysteine. The mutant peptide pPorA-K24C formed an octameric pore that proved to be cation-selective and blocked by cationic α-cyclodextrins. Overall, this study highlighted the power of engineering synthetic peptides to form transmembrane pores of different sizes and ion selectivities. Solid-phase synthesis can provide peptides containing unnatural amino acids, which could allow one to probe chemical reactions and molecular recognition inside the lumen of a single pore.
The generalized transport reaction catalyzed by PorA is:
small molecules (out) → small molecules (in).